I am not an expert on LSD. I do not know how to make LSD. Even if I know, I probably wouldn't tell you. I published this paper while earning a masters in bioengineering at the University of Utah. All of the information found in this report is available on the Internet or your local medical library. Please do not pick up your phone and call me with questions. That is just plain creepy. If you think that I'm a bit negative, it is because I still receive occasional mail like this:
You can learn more about me by exploring the menu located at the top of this page.WE FROM ISRAEL :-) WE LOVE TRIPS WE LOVE GRASS WE LOVE LSD XTC .CAN I BUY IT FROM YOU ?
If you want, feel free to send me comments and/or suggestions either privately.
LSD is considered to be one of, if not the, most potent hallucinogenic drug known. Small doses of LSD (1/2 - 2 ug/kg body weight) result in a number of system wide effects that could be classified into somatic, psychological, cognitive, and perceptual categories. These effects can last between 5 and 14 hours.
Table 1: Effects of LSD
1,
2,
3
| Somatic | Psychological | Cognitive | Perceptual |
|---|---|---|---|
| mydriasis | hallucinations | disturbed thought processes | increased stimulus from environment |
| hyperglycemia | depersonalization | difficulty expressing thoughts | changes in shape/color |
| hyperthermia | reliving of repressed memories | impairment of reasoning | synaesthesia (running together of sensory modalities) |
| piloerection | mood swings (related to set and setting) | impairment of memory - esp. integration of short -> long term | disturbed perception of time |
| vomiting | euphoria | ||
| lachrymation | megalomania | ||
| hypotension | schizophrenic-like state | ||
| respiratory effects are stimulated at low doses and depressed at higher doses | reduced "defenses", subject to "power of suggestion" | ||
| brachycardia |
The study of hallucinogens such as LSD is fundamental to the neurosciences. Science thrives on mystery and contradiction; indeed without these it stagnates. The pronounced effects that hallucinogens have throughout the nervous system have served as potent demonstrations of difficult to explain behavior. The attempts to unravel the mechanisms of hallucinogens are closely tied to basic research in the physiology of neuroreceptors, neurotransmitters, neural structures, and their relation to behavior. This paper will first examine the relationship between neural activity and behavior. It will then discuss some of the neural populations and neurotransmitters that are believed to by effected by LSD. The paper will conclude with a more detailed discussion of possible ways that LSD can effect the neurotransmitter receptors which are probably ultimately responsible for its LSD.
A Brief Foray Into Philosophy and the Cognitive Sciences
Modern physics is divided by two descriptions of the universe:
the theory of relativity and quantum mechanics. Many physicists
have faith that at some point a "Grand Unified Theory"
will be developed which will provide a unified description of
the universe from subatomic particles to the movement of the planets.
Like in physics, the cognitive sciences can describe the brain
at different levels of abstraction. For example, neurobiologists
study brain function at the level of neurons while psychologists
look for the laws describing behavior and cognitive mechanisms.
Also like in physics, many in these fields believe that it is
possible that one day we will be able to understand complicated
behaviors in terms of neuronal mechanisms. Others believe that
this unification isn't possible even in theory because there is
some metaphysical quality to consciousness that transcends neural
firing patterns. Even if consciousness can't be described by
a "Grand Unified Theory" of the cognitive sciences,
it is apparent that many of our cognitive mechanisms and behaviors
can.
While research on the level of neurons and psychological mechanisms is fairly well developed, the area in between these is rather murky. Some progress has been made however. Cognitive scientists have been able to associate mechanisms with areas of the brain and have also been able to describe the effects on these systems by various neurotransmitters. For example, disruption of hippocampal activity has been found to result in a deficiency in consolidating short term to long term memory. Cognitive disorders such as Parkinson's disease can be traced to problems in dopaminergic pathways. Serotonin has been implicated in the etiology of various CNS disorders including depression, obsessive-compulsive behavior, schizophrenia, and nausea. It is also known to effect the cardiovascular and thermoregulatory systems as well as cognitive abilities such as learning and memory.
The lack of knowledge in the middle ground between neurobiology and psychology makes a description of the mechanisms of hallucinogens necessarily coarse. The following section will explore the possible mechanisms of LSD in a holistic yet coarse manner. Ensuing sections will concentrate on the more developed studies of the mechanisms on a neuronal level.
The Suspects
Researchers have attempted to identify the mechanism of LSD through
three different approaches: comparing the effects of LSD with
the behavioral interactions already identified with neuotransmitters,
chemically determining which neurotransmitters and receptors LSD
interacts with, and identifying regions of the brain that could
be responsible for the wide variety of effects listed in Table 1.
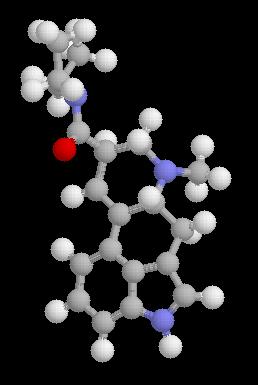
Initial research found that LSD structurally resembled serotonin (5-HT). As described in the previous section, 5-HT is implicated in the regulation of many systems known to be effected by LSD. This evidence indicates that many of the effects of LSD are through
|
| Figure 1: Structure of LSD |
Only a relative handful of neurons (numbering in the 1000s) are serotonergic (i.e. release 5-HT). Most of these neurons are clustered in the brainstem. Some parts of the brainstem have the interesting property of containing relatively few neurons that function as the predominant provider of a particular neurotransmitter to most of the brain. For example, while there are only a few thousand serotonergic cells in the Raphe Nuclei, they make up the majority of serotonergic cells in the brain. Their axons innervate almost all areas of the brain. The possibility for small neuron populations to have such systemic effects makes the brain stem a likely site for hallucinogenic mechanisms.
Two areas of the brainstem that are thought to be involved in LSD's pathway are the Locus Coeruleus (LC) and the Raphe Nuclei. The LC is a small cluster of norepinephrine containing neurons in the pons beneath the 4th ventricle. The LC is responsible for the majority of norepinephrine neuronal input in most brain regions.4 It has axons which extend to a number of sites including the cerebellum, thalamus, hypothalamus, cerebral cortex, and hippocampus.
A single LC neuron can effect a large target area. Stimulation of LC neurons results in a number of different effects depending on the post-synaptic cell. For example, stimulation of hippocampal pyramidal cells with norepinephrine results in an increase in post-synaptic activity. The LC is part of the ascending reticular activating system which is known to be involved in the regulation of attention, arousal, and the sleep-wake cycle. Electrical stimulation of the LC in rats results in hyper-responsive reactions to stimuli (visual, auditory, tactile, etc.)5 LSD has been found to enhance the reactivity of the LC to sensory stimulations. However, LSD was not found to enhance the sensitivity of LC neurons to acteylcholine, glutamate, or substance P.6 Furthermore, application of LSD to the LC does not by itself cause spontaneous neural firing. While many of the effects of LSD can be described by its effects on the LC, it is apparent that LSD's effects on the LC are indirect.4
While norepinephrine activity throughout the brain is mainly mediated by the LC, the majority of serotonergic neurons are located in the Raphe Nuclei (RN). The RN is located in the middle of the brainstem from the midbrain to the medulla. It innervates the spinal cord where it is involved in the regulation of pain. Like the LC, the RN innervates wide areas of the brain. Along with the LC, the RN is part of the ascending reticular activating system. 5-HT inhibits ascending traffic in the reticular system; perhaps protecting the brain from sensory overload. Post-synaptic 5-HT receptors in the visual areas are also believed to be inhibitory. Thus, it is apparent that an interruption of 5-HT activity would result in disinhibition, and therefore excitation, of various sensory modalities.
Current thought is that the mechanism of LSD is related to the regulation of 5-HT activity in the RN. However, the RN is also influenced by GABAergic, catecholamergic, and histamergic neurons. LSD has been shown to also have affinities for many of these receptors. Thus it is possible that some of its effects may be mediated through other pathways. Current research however has focused on the effects of LSD on 5-HT activity. Before specific mechanisms and theories are discussed, a brief discussion of the principles of synaptic transmission will be given.
Overview of Synaptic Transmission
There are two types of synapses between neurons: chemical and
electrical. Chemical synapses are more common and are the type
discussed in this paper. When an action potential (AP) travels
down a pre-synaptic cell, vesicles containing neurotransmitter
are released into the synapse (exocytosis) where they effect receptors
on the post synaptic cell. Synaptic activity can be terminated
through reuptake of the neurotransmitter to the pre-synaptic cell,
the presence of enzymes which inactivate the transmitter (metabolism),
or simple diffusion.
A pre-synaptic neuron can act on the post-synaptic neuron through direct or indirect pathways. In a direct pathway, the post-synaptic receptor is also an ion channel. The binding of a neurotransmitter to its receptor on the post-synaptic cell directly modifies the activity of the channel. Neurotransmitters can have excitatory or inhibitory effects. If a neurotransmitter is excitatory, it binds to a ligand activated channel in the post-synaptic cell resulting in a change in membrane permeability to ions such as Na+ or K+ resulting in a depolarization which therefore brings the post-synaptic cell closer to threshold. Inhibitory neurotransmitters can work post-synaptically by modifying the membrane permeability of the post-synaptic cell to anions such as Cl- which results in hyperpolarization.
Many neurotransmitters that have system-wide effects such as epinephrine (adrenaline), norepinephrine (noradrenaline), and 5-HT work by an indirect pathway. In an indirect pathway, the post-synaptic receptor acts on an ion channel through indirect means such as a secondary messenger system. Many indirect receptors such as muscarinic, Ach, and 5-HT involve the use of G proteins.5 Indirect mechanisms often will alter the behavior of a neuron without effecting its resting potential.
For example, norepinephrine blocks slow Ca activated K channels in the rat hippocampal pyramidal cells. Normally, Ca influx eventually causes the K channels to open. This causes a prolonged after hyperpolarization which extends the refractory period of the neuron. Therefore, by blocking the K channels, the prolonged after hyperpolarization is inhibited which results in the neuron firing more APs for a given excitatory input.5
Other indirect means of neuromodulation include interfering with pre-synaptic neurotransmitter synthesis, storage, release, or reuptake. Inhibiting the reuptake of a neurotransmitter, for example, can cause an excitatory response. Stimulation of neurotransmitter receptors can have a variety of effects on both pre and post-synaptic cells. Pre-synaptic receptors are sometimes involved in self regulation while post-synaptic receptors can cause an increase (excitation) or decrease (inhibition) of AP firing in a neuron. A subtler method of neuromodulation involves molecules that effect these neuroreceptors. Molecules that excite a receptor are referred to as agonists while those that interfere with receptor binding are called antagonists. For example, 5-HT often acts as an inhibitory neurotransmitter. A 5-HT receptor antagonist could interfere with the activation of post-synaptic 5-HT receptors causing them to be less responsive to inhibition. This disinhibition would make the post-synaptic cell more responsive to neural inputs, most likely resulting in an excitatory response.
Theory: LSD Pre-synaptically Inhibits 5-HT Neurons
Raphe Nuclei neurons are autoreactive; that is they exhibit a
regular spontaneous firing rate that is not triggered by an external
AP. Evidence for this comes from the observation that RN neural
firing is relatively unaffected by transections isolating it from
the forebrain. Removal of Ca++ ions, which should block synaptic
transmission, also has little effect on the rhythmic firing pattern.
This firing pattern however is susceptible to neuromodulation
by a number of transmitters.7
In 1968, Aghajanian and colleagues observed that systemic administration of LSD inhibited spontaneous firing of these autoreactive serotonergic neurons in the RN. Serotonergic neurons are known to have a negative feedback pathway through autoreceptors (receptors on the pre-synaptic cell that respond to the neurotransmitter released by the cell). This means that an increase in 5-HT levels causes a decrease in the activity of serotonergic neurons. Serotonergic neurons are also known to make synaptic connections with other RN neurons. This could have the result of spreading out the effects of negative feedback to other RN neurons. This led to the theory that LSD causes a depletion of 5-HT through negative feedback in pre-synaptic autoreceptors.7 The depletion of 5-HT was thought to be responsible for the effects on the previously described systems innervated by the serotonergic neurons. A number of subsequent observations have called this theory into doubt however.
While LSD does cause a decrease in the autoreactive firing of RN neurons, this appears to be an effect and not the cause. These observations are considered however to be compatible with a post-synaptic model. Subsequent research found that LSD and other hallucinogens have a high affinity for post-synaptic 5-HT1 and 5-HT2 receptors. In fact there is significant correlation between the affinity of a hallucinogen for these receptors and its human potency. While it seems logical that 5-HT activity is modulated at 5-HT receptor sites, it is possible that LSD could be affecting 5-HT receptor activity indirectly through adrenic or dopaminic pathways. However, blocking these receptors caused no change in LSD's activity on the 5-HT receptors, thus it appears that 5-HT activity is indeed modified by 5-HT receptors.10 While evidence indicates that LSD is a 5-HT1 agonist, it is debated whether the effects on 5-HT2 receptors is agonistic or antagonistic.11
Theory: LSD Post-synaptically Antagonizes 5-HT2 Receptors
Initial post-synaptic theories postulated that LSD was a 5-HT2
agonist. Pierce and Peroutka (P&P), however, argued that LSD
has a number of antagonistic properties and called into doubt
some of the evidence presented as being compatible with agonist
activity. The primary evidence for agonistic behavior comes from
observations that the effects of LSD are inhibited by 5-HT2
antagonists. P&P pointed out that this is not always the
![[LSD & 5HT2 Receptor]](lsd-rec.jpg) |
| Figure 2: LSD Binding at 5-HT2 Receptor |
5-HT2 receptors are connected to a phosphatidylinositol (PI) second messenger system. PI turnover has been found to be stimulated by 5-HT and antagonized by 5-HT2 antagonists. P&P found that nM concentrations of LSD do not stimulate PI turnover. Therefore, LSD does not act as a classic agonist. They also found that nM concentrations of LSD inhibited the stimulatory effect of 10M 5-HT. The ability of LSD to inhibit a concentration 1000x greater is consistent with it being a 5-HT2 antagonist
P&P also point out that the excitatory effects of 5-HT on CNS neurons appears to be caused by a decrease in K+ conductance attributable to activation of 5-HT2 receptors. P&P found that LSD inhibits this effect in rat somatosensory pyramidal neurons. This also is evidence that LSD acts in an antagonistic role.9
The final line of evidence presented by P&P was from smooth muscle studies. The guinea pig trachea contracts when M concentrations of 5-HT are present. The ability of 5-HT antagonists to inhibit this effect correlates with the antagonists affinity for the 5-HT2 binding site. Thus it appears that this muscle contraction is 5-HT2 mediated. It was found that nM concentrations of LSD did not cause muscle contraction and inhibited the agonistic effects of M concentrations of 5-HT. This also is compatible with the actions of an antagonist.
Theory: LSD Post-synaptically Partially Agonizes 5-HT Receptors
Many of the apparent contradictions in evidence in the debate
over whether LSD acts as a 5-HT2 agonist or antagonist
can be reconciled by the theory that LSD acts as a partial 5-HT2
agonist. Dr. Glennon presented a number of arguments for this
theory including data from his own research and from the studies
discussed by P&P in the previous section.
One of the primary tools used by Glennon to determine the effects of various chemicals on the interactions between LSD and 5-HT was drug discrimination training in rats. Rats were trained to discriminate 1-(2,5-dimethoxy-4-methylphenyl)-2-aminopropane (DOM) from saline. Training with DOM stimuli generalized to many indolealkylamine and phenalkylamine hallucinogens. DOM was chosen instead of LSD as a training drug because of concern that LSD had a number of pharmacological effects. It was thought that if the rat was trained with LSD, it might makes discriminations based on one of the pharmacological effects of LSD other than its effects on 5-HT. With this tool, Glennon demonstrated that a number of 5-HT2 antagonists inhibited the ability of rats to discriminate LSD from saline. This indicates that LSD acts as a 5-HT2 agonist. Glennon offered no explanation for P&P's observation that some antagonists such as spiperone do not have this effect. However, spiperone and a few other similar antagonists appear to only be about 40% effective in inhibiting 5-HT2 sites due to its relative nonselectivity.13
As discussed in the previous section, PI turnover has been found to be stimulated by 5-HT and is antagonized by 5-HT2 antagonists. In another study of the effects of LSD on PI turnover, it was found that LSD acted as a partial agonist (it produces approximately 25% of the effect caused by 5-HT). The apparent difference between this second study and P&P's is that the second study tested the effects at a variety of doses. From this it was concluded that while LSD has a higher affinity for 5-HT receptors than 5-HT does, it has a lower efficacy. This is compatible with P&P's observation that nM concentrations of LSD inhibited the stimulatory effects of uM 5-HT. If LSD acted as a partial agonist with low efficacy, it could compete with 5-HT in binding to 5-HT2 receptors. Since 5-HT is a more potent agonist than the LSD, the effects of LSD would appear antagonistic.
Glennon argued that the guinea pig trachea may not be a good example since 5-HT does not work through a PI mechanism in this case. In the rat aorta, however, 5-HT does hydrolize PI and the contractile effects of 5-HT are antagonized by ketanserin (a 5-HT2 antagonist). While LSD was not tested, another hallucinogen, DOB, was found to have an agonistic effect that could be antagonized by ketanserin. This suggests that LSD acts agonistically in the rat aorta. Glennon points out that it may well be the case that in other cases, the effects may be antagonistic. However, these effects could be explained if LSD had a low efficacy for the receptor.
Hyperthermia and platelet aggregation are both affected by 5-HT2 mechanisms. Hallucinogens such as LSD have been shown to behave agonistically and in the case of platelets, to be antagonized by 5-HT2 antagonists such as ketanserin.11
LSD often has a biphasic response in which low doses have the opposite effects of higher doses. The head twitch response in rodents is believed to be 5-HT2 mediated. At low doses, it has been found that LSD elicits a head-twitch response while at higher doses it antagonizes the response. The rat startle reflex is amplified at low dosages of LSD while decreased at higher doses. This biphasic behavior can also be explained if LSD behaves as a partial agonist.11
In summary, this theory claims that: "LSD is a high-affinity, low efficacy, nonselective 5-HT agonist; in the absence of another agonist it may function as an agonist, whereas in the presence of a high efficacy agonist, it will function as an antagonist." 11
Theory: LSD Post-synaptically Agonizes 5-HT1 Receptors
Glennon also gave another possible explanation for the antagonistic
activity of LSD. There is some evidence that 5-HT1
receptors have an antagonistic relationship with 5-HT2
receptors. As discussed in the previous section, head twitch behavior
is believed to be 5-HT2 mediated. DOI acts as a 5-HT2
agonist and elicits head twitch. 5-OMe DMT also is a 5-HT agonist
but has less efficacy than DOI. If the subject is pretreated with
5-OMe DMT, the effects of DOI are attenuated (because many of
the receptors are filled with the lower efficacy 5-OMe DMT molecules.)
It has been found that A 5-HT1 agonist (8-OH DPAT)
can also cause DOI attenuation. Other studies have also demonstrated
that 5-HT1 agonists can behave functionally as 5-HT2
antagonists.11
Glennon argued that this theory is lent extra credence from the observation that 5-HT2 and 5-HT1c have similar relationships with various hallucinogens. A number of these hallucinogens have been shown to be 5-HT1c agonists. Like 5-HT2 sites, the affinity of hallucinogens for 5-HT1c sites correlates with their hallucinogenic potency in humans. Thus another explanation of the biphasic behavior of LSD is that increasingly higher doses of LSD cause increased antagonism of the 5HT2 receptor through agonism of 5HT1 receptors.
Although, the pre-synaptic theory seems to be fairly well discredited, it is interesting to note that there is debate as to whether pre-synaptic serotonin autoreceptors are of the 5-HT1 type. Whether serotonergic autoreceptors are 5-HT1 or not, it has been demonstrated that there are also post-synaptic 5HT-1 receptors.12 While the role of these receptors is not completely known, some researchers have hypothesized that 5-HT1 receptors may be involved in the regulation of norepinephrine.13 As discussed previously, the majority of norepinephrine neurons are located in the LC which also has system wide innervation.
Recent research on 5-HT receptors calls the theory that 5-HT1 agonism results in 5-HT2 antagonism into question. Since Glennon's paper, the 5-HT1c receptor has been reclassified as 5-HT2c. Since the 5-HT2 receptors discussed in this paper belong to the same family as what was called the 5-HT1c receptor, these have been reclassified as 5-HT2a.14 Since "5-HT1c" is a member of the 5-HT2 family, it is not surprising the LSD affinities are similar for the two receptors. While these reclassifications do not necessarily discount the theory that one receptor has an antagonistic effect on the other, it seems likely that the evidence for this may need to be re-evaluated in terms of recent findings.
Conclusion
The lack of understanding about the mechanisms of LSD is indicative
of the problems involved in the bridging of the worlds of psychology
and neurobiology. As more is learned about the roles and interactions
of various neurotransmitters, receptors, and on a larger scale:
portions of the brain, the mystery will be further unraveled.
With this caveat emptor firmly in mind, it seems that the
best explanation of LSD's effects is that it behaves as a high
affinity partial 5-HT agonist. Depending on the presence of other
molecules and its own concentration, LSD can have either agonistic
or antagonistic effects on post-synaptic 5-HT2 family
receptors. This modulation of 5-HT behavior is probably responsible
for many of the effects attributable to LSD. LSD also has an affinity
for other neurotransmitter receptors that play important roles
in the brain stem such as norepinephrine, dopamine, and histamine.
It is also hypothesized that LSD may modulate neural responses
to these transmitters through its activity on 5-HT1
receptors. Both the Locus Coeruleus and the Raphe Nuclei are
part of the ascending reticular activating system which is implicated
in the sensory modalities. The inhibition of 5-HT in the RN and
release of norepinephrine from LC neurons results in a flood of
information from the sensory system reaching the brain. Some of
the cognitive effects of LSD could be attributed to the effects
of brain stem innervation to areas of the brain such as the cerebral
cortex and the hippocampus.